|
1
|
- Presentation by:
- Vincent E. Giuliano, Ph.D.
|
|
2
|
- In advanced countries, human longevity has been increasing for
millennia. Over the last 100
years, average lifespan from birth in the US increased from 55 to over
79 now.
- Lifespan and health are mostly questions of epigenetics which can evolve
rapidly, not genetics.
- Attention to lifestyle and diet can likely increase human life
expectation by 10 years or more – and lack of attention can decrease it
by a lot more than that.
Knowledge of Nrf2 is an important newer factor.
- Substances in the advanced research pipeline like resveratrol and
rapamycin analogs possibly could further increase our average life expectation by another 7 years
|
|
3
|
- Maximum and average lifespans of simple organisms like nematodes can be
increased by a factor of seven
- The maximum established lifespan of humans is 122
- Key questions of concern to me are:
- What are the prospects for breaking through the limit to allow
lifespans of hundreds of years?
- Given what is known now, how is that likely to come about?
- And, when?
- This presentation addresses these questions.
|
|
4
|
- We all know what aging is, a process through we progressively become
more vulnerable to diseases and disabilities and eventually die. Aging is growing older.
- There are many special and candidate theories regarding the causes of
aging, each with its own group of proponents, and each based on credible
research evidence.
- Each such aging theory is valid in its own domain but explains only a
small part of the picture.
- The author has studied 14 such theories and a number of additional recently-identified candidate theories
- Twenty of these special and candidate theories of aging are elaborated
in the author’s treatise
- ANTI-AGING FIREWALLS –
THE SCIENCE AND
TECHNOLOGY OF AGING
- www.vincegiuliano.name/Antiagingfirewalls.htm
|
|
5
|
- Oxidative damage
- The most traditional theory: that aging is due to accumulated tissue
damage due to oxidative stress created by free radicals.
- 2. Cell DNA damage
- That aging is due to accumulated damage in cellular DNA, leading to
cancers, cell senescence or cell death, in turn leading to tissue and organ
deterioration.
- 3. Mitochondrial damage
- Mitochondria (energy-producing organells in cells) are critical to the
cell reproduction cycle and their DNA, different from the cell’s main
DNA, is particularly vulnerable to damage.
- 4. Tissue glycation
- With aging, tissues become increasingly damaged and dysfunctional due
to cross-linkages with sugar molecules.
- 5. Lipofuscin accumulation
- Metabolic product gunk called lipofuscin accumulates in cells and
inhibits their functionality.
- 6 . Chronic Inflammation
- Chronic inflammation appears to be a core condition underlying many if
not most age-related disease processes.
|
|
6
|
|
|
7
|
- A simplified example is that Oxidative Damage is not only responsible
for Cell DNA Damage and but it also activates NF-kappaB (the Programmed
Epigenomic Changes theory of aging) resulting in expression of
pro-inflammatory genes leading to a Chronic inflammatory response.
- The inflammatory response is a mechanism in turn deeply implicated in Susceptibility
to Cancers and Susceptibility to Cardiovascular Diseases, and Neurological
Degeneration.
- Oxidative Damage also leads to multiple other negative conditions
including Cell DNA Damage and Mitochondrial Damage. Nothing is really
simple, however. Whether a cancer cell is subject to apoptosis or
proliferates is dependent on other Epigenomic considerations such as the
availability of a strong P16 or P53 defense.
|
|
8
|
- The feedback loops between many body systems are so tightly interrelated
that it is difficult to say where one theory leaves off and another one
starts. Examples:
- Neurological degeneration could be triggered by oxidative damage,
lipofuscin accumulation, inflammation, faulty mitochondrial signaling or
defects in energy production.
- Chronic Inflammation is a part of an aging-related program that can be
triggered by numerous stimuli, is generally mediated by a gene
activation sequence triggered by overexpression of NF-kappaB and
related factors, and is an entry portal to several other of the aging
related conditions including Tissue Glycation, Susceptibility to
Cardiovascular Disease, Neurological Degeneration, and Susceptibility
to Cancers.
- Telomere Shortening and Damage is one possible cause of cell
senescence, apoptosis and mutations which in turn generate a number of
the epigenomic-mediated aging conditions including Immune System
Deterioriation, Susceptibility To Cancers, Susceptibility to
Cardiovascular Disease, Neurological Degeneration, and atrophy of
hormone-producing organs leading to Declines in Hormone Levels.
- The later is an example of the feedback between cell-level and
organ-level functioning showing how damage on one level can cause
damage on the other level as well.
|
|
9
|
- I suggest that two of the aging theories provide overall perspectives on
which a SYSTEMS THEORY OF AGING could possibly be built. These are:
- Programmed epigenomic changes
- Decline in functioning of the stem cell supply chain
- These two “framework” theories have four important properties:
- They are elegant and simple.
- They explain how the other theories of aging fit in and the underlying
mechanisms that are associated with them.
- They are applicable across a wide variety of species, including mammals.
- In fact, they themselves are two compatible ways of looking at aging
from contemporary molecular biology and cell biology viewpoints.
|
|
10
|
- In its most basic form this theory holds that aging is not just the
result of accumulation of damage, as many of the aforementioned theories
do, but is the result of some generalized kind of program that unfolds
through life from conception to death. This is a relatively old concept
based on evolutionary arguments, but one with strong new supporting
evidence.
- The emerging concept is that hundreds of genes are involved in what we
call aging, and that there is one or several master programs according
to which these genes are switched on and off through a lifetime in an
intricate pattern to produce early growth, maturation and, finally, assured
death.
- Cellular DNA and RNA mutational damage accumulates stochastically and is
an essential contributor to the aging program.
- Epigenomics provides a general framework for explaining aging as a
programmed phenomenon. Epigenetics is concerned with both heritable and
non-heritable changes in gene expression and activity and also stable,
long-term alterations in the gene transcriptional potential of a cell.
|
|
11
|
- Epigenetic information is based on the experience of a cell, is stored
mainly via DNA methylation,
histone acetylation, non-coding RNA and protein folding patterns,
may be passed on in the process of cell division, and may be accumulated
over the lifetimes of a cell and all of its ancestors. In addition,
nucleosome repositioning, higher order chromatin remodeling, and
accumulated damage to DNA repair machinery, appear to be involved .
- The epigenomic profiles of cells in an organism changes continuously
over the lifetime of that organism and that set of changes defines what
we call aging. I have outlined three current chains of research that
partially support the idea of programmed epigenomic changes of aging
leading to death.
- Further, epigenetic information may be selectively inherited from
generation to generation. Epigenetic patterns capture ancestral history
of acell that is not in the genes themselves and is unique to every
cell. Changing epigenetic information can drastically alter the nature
and lifespans of organisms and is responsible for much of
evolution. Working much faster
than genetic evolution, drastic changes can happen in a few
generations. Animals can grow
bigger or smaller and change their shapes in response to changed
environmental conditions
|
|
12
|
- Gene promoter-region DNA methylation
- An important mechanism for storage of epigenetic information is DNA
methylation, a process by means
of which sites adjacent to genes on chromosomes (promoter regions) are
chemically methylated after a cycle of DNA replication. (Addition of a methyl group to the 5
position of the cytosine ring in a cytosine guanine basepair (CpG))
- Methylation is carried out by DNA methyltransferases using
S-adenosyl-methionine (SAM) as a methyl group donor.
- Methylation is inheritable - passed on in the course of cell divisions
and through generations of people.
- Methylation generally suppresses gene expression by
- physically impeding transcription of proteins. or
- recruiting other CpG binding proteins that change histone structure.
- Occurs in 60-90% of genes.
- Transcriptional silencing due to gene methylation is a central action
required for a number of basic biological processes including embryonic
development, protection against intragenomic parasites, X-inactivation,
genomic imprinting and cognitive functions.
|
|
13
|
- Gene promoter-region DNA methylation (continued)
- At GpG islands containing regulatory genes, unmethylated DNA may become
hypermethylated leading to aberrant conditions like non-small cell lung
cancer and acute lymphoblastic leukemia.
- Aberrant DNA methylation is implicated in multiple disease processes including
atherosclerosis, helicobacter pylori infection and myelodysplastic
syndromes.
- Inappropriate silencing of tumor suppressor genes like P53 and P21
because of aberrant promoter methylation has recently been identified
as a major cause leading to cancer.
- Silencing of “junk DNA,” repetitive sequences between genes, is thought
to be a major biological role of DNA methylation. As it turns out most of the junk is
far from junk
- Abnormal methylation also
predominantly occurs at repetitive sequences, meaning
herterochromatin are affected most.
(inactive genes become active)
- Methylation patterns can regulate genes throughout life.
- The DNA methylation profiles of individuals are unique, change with
aging, and include valuable clues to disease and treatment
progress. For example, DNA methylation of tumor suppressor genes
predicts the relapse risk in acute myeloid leukemia for patients in
clinical remission.
|
|
14
|
- Aberrant methylation may be due to multiple macroscopic causes
- Aging
- Diet : what people eat now, what they ate in the past and what their
parents ate
- Lifestyle factors like exercise
and sleep patterns
- Inflammation, carcinogens, and diseases are known to cause methylation
alterations.
- Tobacco, alcohol, arsenic, and asbestos are associated with
methylation-induced-gene-inactivation.
- Hypermethylation of tumor suppressor genes is found in lung tissue of
smokers.
- DNA methylation is being intensely studied. The database Pubmed.org shows 35,766
research publications related to this topic.
|
|
15
|
- Histone acetylation
- Histone acetylation, another important mechanism of epigenomic change,
relates to folding of histones, the protein spindles around which DNA
is wrapped. Patterns of histone
acetylation are also part of epigenomic memory.
- Histones regulate gene expression by affecting the structure of
chromatin and thus affect the ability of transcriptional activators
and repressors to access regulatory DNA sequences. i.e. an unfolded (acetylated)
histone provides more physical access to a gene promoter sequence than
does a folded (deacetylated) histone.
- Acetylation can be caused by stress, heat-shock factors, acetyltransferases
(HATs). Deacetylation is caused
by histone deacetylases (HDACs).
- Thus, via relaxing chromatin structure, gene expression is stimulated
by HATs which allow transcription factors access to get to DNA . On the other hand, deacetylation of
histones by HDACs promotes chromatin condensation and represses
gene expression. Histone
deacetylation, like DNA methylation, can result in gene silencing.
- Posttranslational modifications of histones is thought to
be an important part of the epigenetic "code"
that determines patterns of cellular gene expression.
- The mTOR “longevity” signaling
pathway is affected in normal and cancer cells by HATs.
|
|
16
|
- Histone acetylation (continued)
- Acetylation/deacetylation are
known to be involved in the activation or silencing of multiple
“aging,” “longevity” and stress-regulating genes including FOXO4,
SIRT1.
- Longevity proteins SIR2 and SIRT1 promote global deacetylation of
histones. It is thought that
this deacetylase activity is responsible for silencing, recombination
suppression and extension of life span in vivo in lower organisms.
FOXO4 is deacetylated by SIRT1 promoting stress-regulating genes and
cellular survival
- A main mechanism used by curcumin, resveratrol and other dietary
polyphenols for inhibition of gene activation by NF-kappaB appears to
be histone deacetylation.
Another is activation of Nrf2.
- There can be significant interplay between histone acetylation and DNA
methylation, particularly when
gene silencing is involved.
|
|
17
|
- Lifelong accumulated damage in cellular DNA repair machinery in adult
stem cells appears to be a major cause of stem cell senescence
- “Cellular aging is linked to deficiencies in efficient repair of DNA
double strand breaks and authentic genome maintenance at the
chromatin level. Aging poses a significant threat to adult stem cell
function by triggering persistent DNA damage and ultimately cellular
senescence.”
- Further, “65% of naturally occurring repairable DNA damage in
self-renewing adult stem cells occurs within transposable elements”
of Alu RNA/DNA.
|
|
18
|
- Telomere shortening
- My current view is that
telomere lengths are downstream consequences of other cell state
factors. Telomeres generally
shorten with cell reproduction but can also lengthen depending on
complex feedback loops. Cell
senescence can lead to short telomeres which can contribute to,
apoptosis, or malignant transformation. Cell senescence is largely driven by
other factors than telomere lengths.
- Three years ago, I subscribed to the notion that extending telomere
lengths could be a iife-extending intervention, but no longer do.
- Because rats and mice have long
telomeres throughout life, telomere shortening by itself does not
fully explain mammalian aging.
|
|
19
|
- Increased expression of a nuclear factor NF-kappaB
- expression of NF-kB appears to be one of the body’s regulatory means
for handling situations of stress, cancer, damage and disease. In
eukaryotic cells NF-kB is an important regulator of genes that control
cell proliferation and cell survival.
- NF-kB regulates anti-apoptotic genes that protect healthy cells from
cell death and activates the expression of genes that keep cells
proliferating.
- On the other hand, activated NF-kB binding to genes has long been
known to play a central role in promoting runaway inflammation and
inflammation’s negative consequences.
- Recent studies indicate that
NF-kB signaling appears is
a major regulator of gene expression affecting hundreds of
genes related to the aging progress. NF-kB cell signaling has been
shown to be a meta-factor for determining aging of a number of key
cell types
- Inhibition of NF-kB signaling
is being researched as a
cancer therapy and is thought also to be a possible approach to life
extension
|
|
20
|
- P16/Ink4a, a tumor suppressor gene, becomes increasingly active with
age in mammals. It is a known mediator of cell senescence and biomarker
of aging as well as a possible promoter of mammalian aging. P16/Ink4a
works together with three other genes to articulate a process of
simultaneously protecting against cancers and shutting down adult stem
cell function and regenerative capacity in aging tissues.
- aging-related decline of efficacy of DNA repair machinery might
possibly result from promoter methylation of the Mms22 gene, resulting
in increasing susceptibility to oxidative damage with age.
- Promoter methylation of the P21 and P53 apoptosis genes can result in increased
susceptibility to cancers. HDAC
inhibitors can help them turn back on again.
- The P66Shc gene, associated with longevity in mammals, appears to be
silenced through some combination of deactylation (resulting in protein
folding) and cytosine methylation.
|
|
21
|
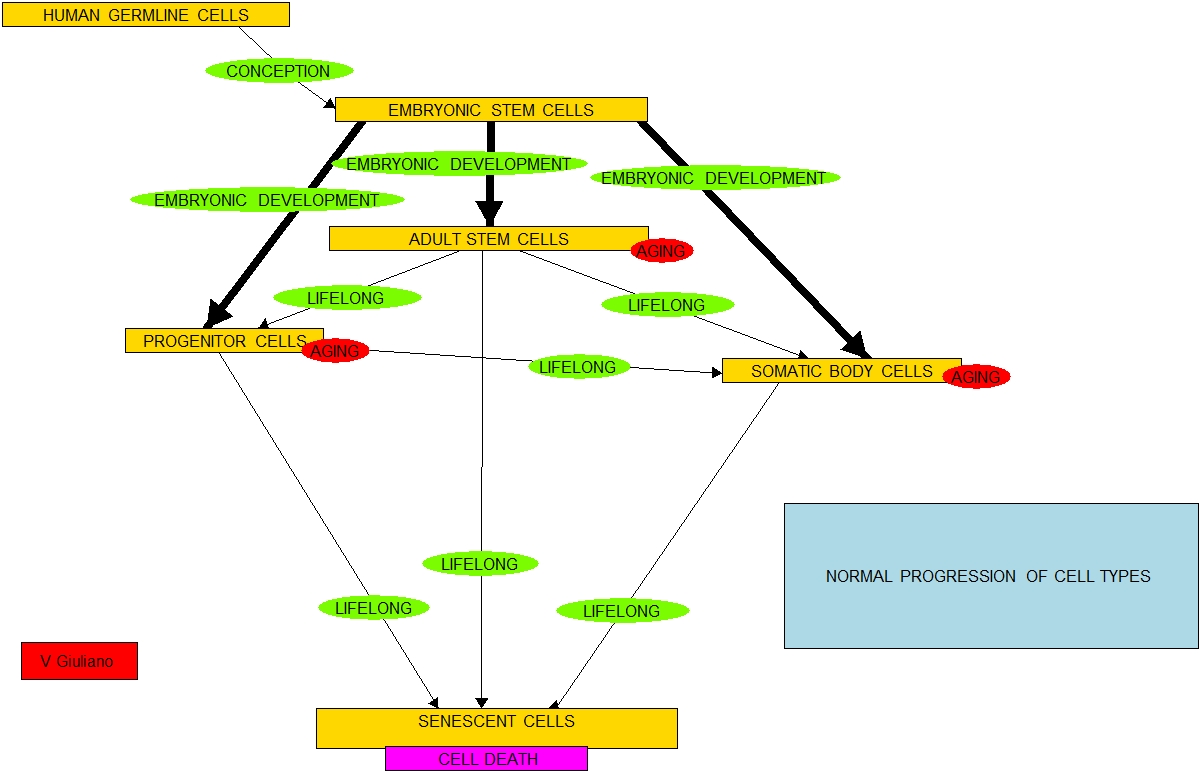
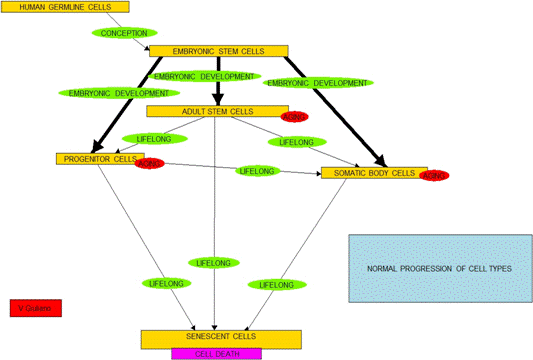
- In a simplified model, think of the 210 kinds of cells found in the
human body as falling in five categories:
- A. Pluripotent cells, ones which
are and capable of differentiating into any other cells. Human
embryonic stem cells (hESCs) and induced pluripotent stem cells
(iPSCs) are in this category,
- B. Relatively undifferentiated multipotent
somatic stem cells, such as may exist in bone marrow or vascular walls
(e.g. hematopoietic stem cells, mesenchymal stem cells and pericytes). These multipotent cells are each
capable of differentiating into a variety of kinds of somatic cells.
- C. More differentiated stem and progenitor
cells (e.g. endothelial progenitor cells, myoblasts or satellite cells
in muscle tissue). These are
cells capable of differentiating only into specific somatic cell types.
- D. Normal body somatic cells
(e.g. cardiomyocytes, red blood
cells, leukocytes, keratinocytes, melanocytes, and Langerhans cells).
- E. Senescent cells, ones which
no longer can divide.
- The list is in order of increasing cell-type specificity and decreasing
potency to differentiate into other cell types. Starting at conception and throughout
life, all cells on this list except the senescent ones will selectively
reproduce and possibly differentiate into cells of types further down in
the list.
- In some cell lines there are actually many more intermediate forms of
progenitor cells, but a model of five categories of cells is sufficient
for this discussion.
|
|
22
|
- Cells in all categories except Type E can divide to make new cells. They are all subject to mutation, cell
damage, oncogenesis and, it is thought by some, are subject to replicative
senescence.
- Cells of Type A in the early embryo progressively differentiate to make
all cells of Types B, C or D.
- All cells of Type D result from differentiation of cells of Type A, B
and/or C, possibly via intermediary progenitor and stem cell types.
- Some cells of Type B may differentiate through several intermediate
forms before creating Type D cells.
Hierarchy in differentiation is always preserved under natural
conditions, although it may or may not necessarily be the case that
intermediate stem cell types are involved depending on the kind of cell.
- An early embryo consists of A-Type cells. This supply-chain process continues
through life although in aging there will be more and more cells of
Types D and E and fewer and fewer active cells of Types B and C. and virtually
no active Type A cells left.
- Healthy normal aging is thus a matter or cellular supply chain
management. The body must assure
that there are not too many Type E cells around for they create havoc.
|
|
23
|
- Type D cells are the workhorses of day-to-day functioning and the key
factors involved are insuring a good supply of them by avoiding
damage-related or replicative senescence, taking care of their need for
nutrition and a healthy intra-cellular environment, and making sure that
damaged or proto-cancerous cells are eliminated through proper
apoptosis. Also, it is important
to assure that Type B and C cells are able to differentiate properly to
provide a reliable continuing source of replacements for the Type D
cells.
- The issues for Types B and C cells include seeing that they are in
sufficient supply and health so as to be able to differentiate into Type
D cells and making sure that the differentiating option is readily
available when needed. Other
issues for Types B and C cells are similar to those for Type D cells -
preventing damage-related or replicative senescence, and preventing
oncogenesis.
- In aged individuals there are few if an active Type A cells around to
replace Type B and C cells as they are lost, a possible major reason for
aging.
|
|
24
|
|
|
25
|
|
|
26
|
- The supply chain mechanisms appears to be operable throughout life in a
manner controlled by several feedback mechanisms. E.g. In the absence of
P21, hematopoietic stem cells would not remain quiescent in their niches
but would instead prematurely differentiate when stress occurs
exhausting the pools of those cells leading to premature death.
- The balance of cell Types is highly dependent on the stage of
development of the organism, favoring gradual shift to the
more-differentiated cell types
- Adult stem cells live in niches -
stem cell microenvironments and the health of the stem cells and their
ability to reproduce or differentiate both depend upon and condition the
states of their niches.
- Health for people of all ages requires continuing operation of the
supply chain throughout life.
But it is a once-through process that runs out of adult stem
cells with old age.
|
|
27
|
- Proliferation and differentiation of Type A, B, and C stem and
progenitor cells decreases with aging.
This appears to be associated with buildup of NF-kappaB and
P16/Ink4a.
- Although the mobilization responsiveness of Type C stem cells declines
with age, it appears that their regenerative capability can to some
extent be restored through environmental messages or induction of Notch
activity.
- The gene expression profiles in
Type A human embryonic stem cells offer regenerative anti-aging
potential not found in more mature stem cells.
- This matter of concern here is that advanced aging is due to a slowing
rate of organ regeneration due to declining SSC differentiation
activity, this in turn being due to exhaustion of pools of Type B and
Type C stem cells because of differentiation and replicative senescence.
|
|
28
|
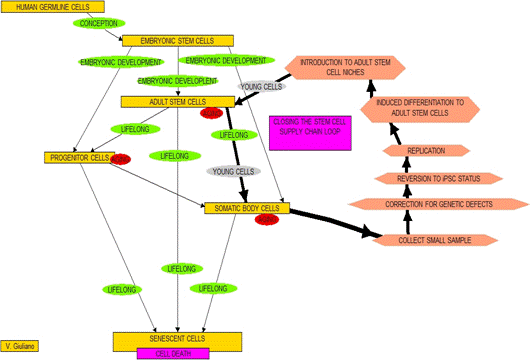
- The stem cell supply chain is now a once-through in life process
- Embryonic stem cells disappear during fetus development
- All other stem cells are subject to replicative senescence and decline
in differentiation capability with age
- Thus with advancing age, normal somatic Type D cells are no longer
replaced and the diseases and problems of aging ensue
- In the longer view there is hope of “closing the loop” in the stem cell
supply chain and making it a continuous process. The concept, for
example, is:
- to revert a person’s skin or blood cells to Type A cells using induced
pluripotent stem cell (iPSC) technology,
- To correct those iPSCs for any
genetic defects using gene splicing,
- To induce those Type A iPSCs to differentiate into Type B and Type C
stem cells, and
- to introduce those adult stem cells back into their niches so they can
continue to exercise their regenerative functions.
|
|
29
|
|
|
30
|
- For this concept to become real a number of technical research
challenges must be met including:
- Obtaining iPSCs that are free of DNA contamination, that have long
telomeres and full hESC pluripotency
- Developing reliable means for assuring differentiation into Types B and
C stem cells
- Developing reliable and safe means for introducing those cells into their respective
body niches
- Much research is being devoted to these approaches but 10-20 years are
likely to be required before the
stem cell supply chain can truly be closed in humans.
- If and as this happens I conjecture that extraordinary human longevity
might become possible – lifespans of hundreds of years.
|
|
31
|
- They are two sides of the same coin, representing differing viewpoints
of molecular biology and cell biology.
- Of course all the cells in an individual have the same genome but
their DNA acquires additional epigenomic markers as they
differentiate. So, looked at in
terms of DNA, the differences between kinds of cells including stem
cells is one of epigenomics,
e.g. DNA methylation, histone acetylation, binding-site molecular folding,
alterations in non-coding RNA, etc..
- Factors that can revert fully mature Type D cells to Type A
pluripotent cells and possibly more differentiated stem cell types are
being actively researched. Many
new combinations of reprogramming factors have been discovered in
addition to the original ones:
(Oct3/4, Sox2, Klf4, cMyc)
|
|
32
|
- They are two sides of the same coin, representing differing viewpoints
of molecular biology and cell biology.
For example: (cont.)
- Several proteins seem to play key roles in stem cell differentiation, survivin,
an apoptosis inhibitor that is a target of cancer therapies, being a
key one. FoxO family members play a critical role in these physiologic
processes in the HSC compartment and thereby regulate maintenance and
integrity of HSCs.
- Effective mTORC1 negative regulation is essential for keeping the
critical balance between stem cell self-renewal and differentiation:
Too little self-renewal or too much differentiation of hematopoietic
stem cells may jeopardize the ability to sustain hematopoiesis
throughout life, whereas excessive self-renewal and/or aberrant
differentiation may result in leukemogenesis.
- TAp63 serves to maintain adult skin stem cells by regulating cellular
senescence and genomic stability, thereby preventing premature tissue
aging
|
|
33
|
- (Lunyak et al, others 2008 - 2012) have established that:
- Stem cell senescence results from aging and demonstrably leads to
diseases and aging.
- Stem cell senescence is an epigenetic phenomenon. In fact, SINE/Alu
Retrotransposons are transcriptionally up-regulated upon Senescence of
hADSC, and this affects chromatin structure and impairs the DNA damage
repair machinery.
- Epigenetic interventions can reverse cell senescence markers affecting
aging
- Small non-coding RNA species (like IncRNAs, shRNAs, siRNAs and piALU
RNAs) can play critical roles in gene regulation, DNA repair and
chromatin regulation
- In fact, SINE/Alu Retrotransposons are transcriptionally up-regulated
upon Senescence of hADSC, and this affects chromatin structure and
impairs the DNA damage repair machinery.
|
|
34
|
- “Cellular aging is linked to deficiencies in efficient repair of DNA
double strand breaks and authentic genome maintenance at the chromatin
level. Aging poses a significant threat to adult stem cell function by
triggering persistent DNA damage and ultimately cellular senescence.”
Further, “65% of naturally occurring repairable DNA damage in
self-renewing adult stem cells occurs within transposable elements” of
Alu RNA/DNA.
- Specific interventions involving removal of damaged specific segments
of RNA, formerly thought to be “junk RNA,” can reverse adult stem cell
senescence. Working with specific segments of RNA can add to the
traditional epigenetic interventions that mainly have related to DNA
methylation and histone acetylation. Specifically, by modifying a
Lentivirus genome to express GFP and sh-RNA against Alu transcript, it
is possible to knock down the generic SINE/Alu transcript in senescent
adult stem cells, reversing senescence markers, rejuvenating the
cells, and restoring their lost differentiation capability.
- So far, this has been achieved in-vitro. It is yet to be shown that it can be
accomplished in-vivo
|
|
35
|
- They are two sides of the same coin, representing differing viewpoints
of molecular biology and cell biology.
For example (continued):
- Buildup of levels of Ink4a/P16 associated with aging slows down the
rate of differentiation of adult stem cells. P16/Ink4a works together
with three other genes (Arf, Hmga2 and let-7b) to articulate a
process of simultaneously protecting against cancers and shutting
down adult stem cell function and regenerative capacity in aging
tissues.
- In young cells, Polycomb group proteins act on the INK4/ARF gene
regulatory domain so as to the keep the expression of P16(INK4a)
turned off, the gene is silenced. In senescent cells, however,
there are epigenetic modifications (DNA and histone methylation
changes) which block the inhibitory actions of the polycomb group
proteins, so the P16(INK4a) and Arf genes are activated. So,
cell senescence leads to another pro-aging effect, the activation of
the P16(INK4a) and Arf genes which in turn slows down stem cell
differentiation
- aging-related decline of efficacy of DNA repair machinery might
possibly result from promoter methylation of the Mms22 gene, resulting
in increasing susceptibility to oxidative damage with age.
- Promoter methylation of the P21 and P53 apoptosis genes can result in increased
susceptibility to cancers.
- Recent research has shown that adequate expression of the P21 gene is
necessary to keep adult stem cells from differentiating prematurely
resulting in exhaustion of their
supply.
- The P66Shc gene, associated with longevity in mammals, appears to be
silenced through some combination of histone deactylation (resulting in
protein folding) and cytosine methylation(ref). The gene regulates mitochondrial
metabolism.
|
|
36
|
- With respect to the accumulated damage theories (Oxidative damage, DNA
damage, Mitochondrial damage, Incorrect protein folding ):
- Such damage will of course occur; it occurs in younger as well as older
people. The difference is that
apoptosis and DNA repair mechanisms work much better in younger people
as is explained by the Programmed Epigenomic Changes theory. Also, dead cells in younger people
are replaced by ready stem cell differentiation but not so in older
people as explained by the Stem Cell Supply Chain theory.
- With respect to inflammation, immune system, susceptibility to cancer,
cardiovascular and neurological disease theories
- There is widespread acknowledgement that epigenomic reprogramming plays
major roles in disease susceptibilities and development in these areas
- There is much current research
on developing HDAC inhibitors as preventative and therapeutic agents
for such diseases
- There is also much research focusing on stem cell therapies in each of
these disease areas and on
aspects of operation of the stem cell supply chain
- With respect to the telomere shortening theory of aging
- Expression of telomerase and other factors affecting telomere length is
determined by the epigenetic state of the cell
- It was once thought that telomere shortening could be a major factor in
the age-related decline of proliferative and differentiation
capabilities of adult stem cells.
Lunyak’s work at the Buck Institute suggests that the smoking
gun is instead damage to the RNA-encoded DNA damage repair machinery in
adult stem cells.
|
|
37
|
- This presentation is available online at
- http://www.vincegiuliano.name/AAAS5-18_files/frame.htm. The blue hyperlinks are active in the
online presentation
- There is far more of relevance
than can be included here.
Details of he theories of aging can be found in my
frequently-updated online treatise:
- ANTI-AGING FIREWALLS – THE SCIENCE AND TECHNOLOGY OF LONGEVITY at
- www.vincegiuliano.name/Antiagingfirewalls.htm
- And many detailed discussions on topics raised in this presentation can
be found in my BLOG:
- www.agingsciences.com
- Vincent E. Giuliano, Ph.D. vegiuliano@comcast.net
|

 Notes
Notes{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}